Название: Литические взаимодействия сапротрофных бактерий и дрожжей в естественных экосистемах
Вид работы: реферат
Рубрика: Биология и химия
Размер файла: 45.59 Kb
Скачать файл: referat.me-21064.docx
Краткое описание работы: Одним из основных факторов, регулирующих состав и стабильность микробных сообществ в природных экосистемах, является взаимодействие между составляющими их популяциями микроорганизмов.
Литические взаимодействия сапротрофных бактерий и дрожжей в естественных экосистемах
Т. Ф. Черняковская, Т.Г.Добровольская
Одним из основных факторов, регулирующих состав и стабильность микробных сообществ в природных экосистемах, является взаимодействие между составляющими их популяциями микроорганизмов. Взаимодействия могут быть как положительными, так и негативными. Вариантом проявления антагонистических взаимоотношений является выделение литических ферментов микроорганизмами одной группы, воздействующих на клетки микроорганизмов другой группы. Известно, что некоторые бактерии и актиномицеты способны лизировать клетки бактерий, дрожжей и грибной мицелий, так как обладают соответствующим набором ферментов, в том числе глюкозидазами, хитиназами, липазами (Webley, Jones, 1971, Prokaryotes, 1981; Кулаев, 1997). Различают эндогенные (внутриклеточные) литические ферменты, которые находятся в клетках продуцентов в непосредственной близости от объектов своего действия, и экзогенные, то есть секретируемые в среду обитания образующих их организмов (Кулаев, 1997).
Логично предположить, что литиче-ская активность в наибольшей степени может проявляться в тех природных местообитаниях, где разные группы микроорганизмов взаимодействуют в условиях постоянного поступления растительного материала и на разных стадиях его переработки в общем круговороте веществ. К таким местообитаниям относятся лесные и степные подстилки, гумусовые горизонты почв, торфяники, зоомикробные комплексы. Однако имеются лишь единичные исследования, в которых целенаправленно изучались бы в одних и тех же природных биотопах те группы микроорганизмов, между которыми складываются антагонистические взаимоотношения. Так, было показано (Сорокин, Бабьева, 1982), что некоторые виды бацилл и стрептомицетов лизировали клеточные стенки дрожжей, попадающих в почву из подстилок.
Для изучения взаимодействий бактерий и дрожжей нами выбраны в качестве объектов исследования типичные и луговые степи Русской равнины, а также зоомикробные комплексы диплопод, питающихся растительным опадом. Были изучены численность и таксономическая структура блока сапротрофных бактерий и дрожжей в разных ярусах степных биогеоценозов (БГЦ) и в пищевых цепях диплопод (Черняковская и др., 1990; Вино-варова, 1989; Byzov, et al.,1996). Было установлено, что максимальное количество и таксономическое разнообразие бактерий гидролитического комплекса, приуроченных к деструкции растительных остатков, сосредоточено в степном войлоке (ветоши и подстилке), в то время как дрожжи ассоциированы с живыми и отмирающими растениями, в подстилках их количество резко снижается. В зоо-микробных комплексах диплопод как бактерии, так и дрожжи сосредоточены в содержимом заднего отдела кишечника и экскрементах.
В ходе исследования была составлена коллекция бактерий (насчитывающая более 600 штаммов, относящихся к 19 родовым таксонам) по принципу отбора доминирующих групп из каждого компонента биогеоценоза или зоомикробного комплекса. Для проверки литических свойств бактерий были отобраны также штаммы дрожжей разных филогенетических линий из коллекции кафедры биологии почв МГУ и культуры дрожжей, применяемые в промышленности, − Candida maltosa и Saccharomyces cerevisiae.
Методы исследования
Дрожжелитическую активность бактерий определяли на твердой агаризо-ванной среде с дрожжевыми клетками в качестве единственного источника угле- рода. Культуры дрожжей выращивали на среде следующего состава (%): глюкоза -3; (NH4)2SO4 - 5; KH2PO4 - 0, 085; K2HPO4 − 0,015; Mg 2SO4-7HO - 0,05; NaCl - 0,01; CaCl2 - 0,01; дрожжевой ав-толизат - 0,5. Инокулят дрожжей вносили в 200 мл среды, разлитой по колбам, и помещали на качалку. Температура инкубации для различных представителей мезофилов составляла 26−28°, для псих-рофилов - 10°С. Сроки инкубации 2−3 суток или 10−14 суток соответственно. К этому времени культуры дрожжей достигали стационарной фазы. Полученную жидкую культуру дрожжей центрифугировали, промывали дистиллированной водой 2 раза (для удаления внеклеточных полисахаридов)в стерильных условиях. Промытую биомассу смешивали с расплавленным и остуженным до 40−60°С солевым агаром. Состав солевой основы (%): (NH4)2SO4 - 0,1; KH2PO4 - 0,1; Mg 2SO4-7HO - 0,05; агар - 2, рН 7,0. На 500 мл среды брали суспензию клеток из 12 колб, что составляло примерно 0,7 - 1 %-ную концентрацию биомассы дрожжей в среде. Среду с убитыми клетками дрожжей готовили аналогично, но промытую суспензию клеток предварительно нагревали на водяной бане 10 мин при 80°С. 3-х суточные культуры бактерий в виде суспензии наносили на поверхность среды множественным репликатором. Чашки с посевом инкубировали при 24°С 14 суток. Регистрировали зоны просветления, т.е. лизиса суспендированных в толще агара дрожжевых клеток. О степени активности культур судили по размерам зон лизиса вокруг колоний-продуцентов литических ферментов.
Результаты исследований
Способность лизировать клеточные стенки дрожжей была проверена у 628 штаммов бактерий, изолированных из разных субстратов. Среди бактерий, выделенных с зеленых частей растений, а также со стенок пищеварительного тракта диплопод (пристеночное сообщество), доля активных дрожжелитических штаммов незначительна - 5%. Несколько выше их содержание в почве - 18%. Активные штаммы были выделены в основном из подстилок и зоогенных субстратов (содержимого кишечников и экскрементов диплопод). Половина всех выявленных в этих биотопах бактерий проявила дрожжелитическую активность (табл. 1).
Таксономический состав штаммов, составляющих коллекцию и количество активных среди них, представлены в (табл. 2).
Таблица 1
Дрожжелитическая активность бактерий, выделенных из разных субстратов исследуемых биогеоценозов и зоомикробных комплексов
| Субстрат | Выделено и проверено на активность | Из них активны | |
| число | % | ||
| Филлоплана, пристеночное сообщество диплопод | 152 | 8 | 5 |
| Растительные остатки подстилка | 233 | 119 | 51 |
| Содержимое кишечников и экскременты диплопод | 201 | 111 | 55 |
| Почва | 50 | 9 | 18 |
Таблица 2 Дрожжелитическая активность бактерий разных родовых таксонов
| Родовые таксоны бактерий | Количество штаммов | % активных штаммов | |||
| Streptomyces | 40 | 85 | |||
| Promicromonospora | 64 | 81 | |||
| Agromyces | 12 | 75 | |||
| Oerskovia | 111 | 68 | |||
| Bacillus | 56 | 54 | |||
| Mycobacterium | 83 | 42 | |||
| Micrococcus | 30 | 27 | |||
| Cytophaga | 20 | 25 | |||
| Myxococcus | 30 | 17 | |||
| Cellulomonas | 30 | 0 | |||
| Arthrobacter | 30 | 0 | |||
| Rhodococcus | 36 | 0 | |||
| Curtobacterium | 4 | 0 | |||
| Brevibacterium | 2 | 0 | |||
| Pseudomonas | 20 | 0 | |||
| Erwinia | 15 | 0 | |||
| Serratia | 5 | 0 | |||
| Plesiomonas | 20 | 0 | |||
| Vibrio | 20 | 0 | |||
Условные обозначения:0 — литическая активность не выявлена у всех штаммов бактерий
Максимальное количество штаммов, обладающих дрожжелитическими свойствами, было обнаружено среди бактерий родов Streptomyces, Promicromonospora, Oerskovia, Agromyces. Впервые дрожже-литическая активность выявлена у акти-нобактерий родов Mycobacterium и Micrococcus.
Таким образом, способность лизи-ровать клетки дрожжей характерна для бактерий гидролитического комплекса, принимающих активное участие в деструкции растительного опада − Streptomyces, Promicromonospora-Oers-kovia, Bacillus, Mycobacterium, Cytophaga, Myxococcus. Среди обладающих дрож-желитической активностью бактерий больше представителей с грамположи-тельным типом строения клеточной стенки, чем c грамотрицательным.
Чувствительность дрожжей к лити-ческому действию бактерий проявилась как на уровне классов, так и родов. Бактерии лизировали 37% дрожжевых культур, относящихся к базидиомицетам, и 28% штаммов дрожжей - аскомицетов.
Литическое действие бактерий р. Promicromonospora особенно сильно проявлялось в отношении Ph. rhodozyma (типичное местообитание - сокотечения деревьев) - 100% взаимодействий от теоретически возможных; Cr.albidus и Rh. glutinis (типичное местообитание - фил-лосфера растений и подстилки) - 90 и 69% соответственно. Бактерии р. Oerskovia были наиболее активны по отношению к клеткам Rh. glutinis - 65%. Представители р.р. Promicromonospora -Oerskovia проявляли высокую дрожжели-тическую активность и по отношению к почвенным дрожжам L. кononenkoae - 58 и 61% (табл. 3).
Таблица 3 Чувствительность дрожжей разных видов к литическим ферментам бактерий
| Oerskovia | Promicromonospora | Agromyces | Bacillus |
| Debaryomyces vanriji | |||
| Lipomyces kononenkoae | |||
| Lipomyces tetrasporus | |||
| Phaffia rhodozyma |
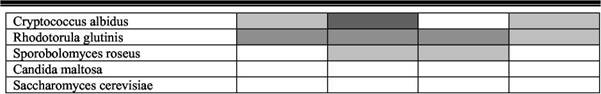
Примечание: Процент штаммов дрожжей, чувствительных к литическому воздействию бактерий.
![]()
Таким образом, в результате проверки коллекции бактерий (более 600 штаммов, представленных 19 родами), выделенных из различных природных биотопов, было установлено, что способность к лизису дрожжей широко распространена среди разных таксономических групп бактерий.
Численность и таксономическое разнообразие дрожжелитических бактерий, среди которых доминировали грам-положительные формы, были максимальны в подстилках и дерновом горизонте почв исследованных биогеоценозов.
Впервые была выявлена способность к лизису дрожжей у представителей родов Micrococcus и Mycobacterium, сосредоточенных на зеленых и отмирающих частях растений.
В зоомикробных комплексах дип-лопод (содержимое кишечника и экскременты), питающихся растительным опадом, обнаружены дрожжелитические бактерии родов Promicromonospora, Oerscovia, Streptomyces, Bacillus, Myxococcus. По-видимому, эпифитные дрожжи, поступающие вместе с растительными остатками в пищеварительный тракт диплопод, подвергаются там утилизации бактериями, имеющими литиче-ские ферменты.
Дрожжелитическая активность бактерий-гидролитиков сапротрофного комплекса подстилок и почв - один из возможных факторов, регулирующих в природных экосистемах состав, численность и соотношение популяций в сук-цессионных процессах разложения растительных остатков. Этот фактор проявляется как при непосредственных взаимодействиях бактериальных и дрожжевых популяций, так и при прохождении растительного материала через пищеварительный тракт беспозвоночных животных, населяющих подстилку и почву.
Список литературы
1. Бабьева И.П. Дрожжи в биогеоценозах разных природных зон // Почвенные организмы как компоненты биогеоценоза. М. :Наука.1984.С.131-141.
2. Виноварова М. Е. Дрожжевые грибы в структуре степных сообществ (заповедные степи Русской равнины): Автореф... дисс. канд. биол. наук. М., 1989. 22 с.
3. Сорокин Д.Ю., Бабьева И.П. Лизис природных популяций дрожжей почвенными микроорганизмами // Микробиология. 1982. Т.51. Вып.2. С. 328-331.
4. Черняковская Т.Ф., Добровольская Т.Г., Лысак Л.В., Ванина С.А. Закономерности распределения эпифит-ных и сапротрофных бактерий по компонентам вертикальной структуры степных биогеоценозов // Почвоведение. 1990. №6. С.68-77.
5. Bysov B.A., Dobrovolskaja T.G., Chernjakovskaja T.F., Zenova G.M. Bacterial communities assosiated with soil diplopods // Pedobiologia. 40. 1996.I
6. The Prokaryotes. A Handbook on Habitats, Isolation and Identification of bacteria: In 2V/Ed. by Starr M.P., Stolp H., Truper H.G., Balows A., Schlegel H.G. - Springer- Verlag etc., 1981. P. 2284.
7. Кулаев И. С. Бактериолитические ферменты микробного происхождения в биологии и медицине // Соро-совский образовательный журнал. 1997. № 3. С. 23 - 28.
Похожие работы
-
Брожение
Брожение, ферментативное расщепление органических веществ, преимущественно углеводов. Может осуществляться в организме животных, растений и мн. микроорганизмов без участия или с участием О2 (соотв. анаэробное или аэробное брожение).
-
Возрастное распределение, выживаемость и смертность виноградной улитки Helix pomatia L. в экосисте-мах Калининградской области
В рамках комплексной работы по изучению экологии и биологии виноградной улитки Helix pomatia на территории Калининградской области проведено исследование возрастного распределения, выживаемости и смертности в трех природных популяциях данного вида.
-
Функции белков
Ниже рассматриваются главные и в некотором смысле уникальные биологические функции белков, несвойственные или лишь частично присущие другим классам биополимеров.
-
Общая характеристика бактерий
Хотя каждая бактериальная клетка очень мала, тем не менее общая масса бактерий, обитающих на Земле, превышает массу всех других живых организмов вместе взятых. Сейчас известно около 2500 видов бактерий.
-
Влияние лактулозы на формирование микробиоценоза пищеварительного канала цыплят-бройлеров
Приводятся результаты исследования по формированию микробиоценоза в зобу, слепых и прямой кишках трех- и семисуточных цыплят-бройлеров, получавших пребиотик с лактулозой. Пребиотик стимулирует заселение просвета кишечника полезной микрофлорой уже в первую неделю жизни цыплят.
-
Инициированные комплексы почвенных грибов в агроценозах
Показано, что различные системы обработки почвы являются мощным антропогенным фактором, нарушающим формирование комплексов грибов, характерных для почв в ненарушенных экосистемах, влияя на их состав и численность.
-
Действие повреждающих факторов на клетку
Действие повреждающих факторов на клетки осуществляется прямо (первичные факторы повреждения) или опосредованно. В последнем случае речь идет о формировании цепи вторичных реакций, реализующих повреждающее влияние.
-
Круговорот веществ и поток энергии в экосистемах
Питание — основной способ движения веществ и энергии. Организмы в экосистеме связаны общностью энергии и питательных веществ, которые необходимы для поддержания жизни.
-
Микробная эндокринология и биополитика
Работа посвящена исследованию эволюционно-консервативных химических факторов, выступающих как гормоны и/или медиаторы у высших животных и человека.
-
Изучение микробных сообществ при повышенном увлажнении в условиях модельного эксперимента
В последние десятилетия одной из наиболее актуальных тем научных исследований в биологии становится изучение биоразнообразия живых организмов, в том числе - прокариот.