Название: Молекулярная эволюция в подсемействе Prunoideae Focke
Вид работы: реферат
Рубрика: Биология и химия
Размер файла: 122.96 Kb
Скачать файл: referat.me-21984.docx
Краткое описание работы: С использованием белковых (молекулярных) маркёров выявлены родственные связи, разработана концепция эволюции внутри подсемейства Prunoideae Focke (Rosaceae Juss.). В результате уточнён возраст родов и видов, центры их происхождения, пути расселения.
Молекулярная эволюция в подсемействе Prunoideae Focke
Молекулярная эволюция в подсемействе Prunoideae Focke
В. И. Авдеев
Крупное подсемейство сливовых (Prunoideae Focke), как и остальные 4 подсемейства Rosaceae Juss., слабо изучены в эволюционном плане, хотя и являются очень ценными растениями. До конца не ясен даже родовой состав подсемейства, генетические связи родов и видов, произрастающих на разных континентах. Биосистематика сливовых, как и большинства других растений, построена на анализе внешних признаков. В этом случае вне поля зрения остаются молекулярные механизмы эволюции, в основе которых лежат явления рецессивности, доминантности, репрессии, дерепрессии генов и др. Поэтому у живых организмов при однородности внешних признаков весьма обычно значительное разнообразие особей, видов, родов по молекулярному составу, биохимическим признакам [1].
Попытки создания системы подсемейства сливовых предпринимались, но они имели чисто практический характер, для обслуживания селекционных нужд [2]. Это были формальные схемы, где не учитывались палеоданные, история флор и ареалов родов, даже полиморфизм популяций по внешним признакам. С учётом всего этого и использованием новейших экспериментальных данных по белковым (полипептидным, молекулярным) маркёрам впервые эволюция подсемейства сливовых была охарактеризована в докторской диссертации и статье [3, 4]. Однако на то время ещё не были уточнены молекулярные критерии эволюционного возраста таксонов, это было сделано уже позднее [5, 6] на основе разработок школы А. В. Благовещенского [7]. В итоге связи по белковым маркёрам внутри родов сливовых и межродовые отношения в процессе их эволюции представлены на дендрограммах (рис. 1 и 2) и разъяснены в описаниях этих рисунков. Краткие же комментарии к этим данным будут нижеследующими.
Древнейшими (верхнемелового возраста) были роды Cerasus Mill. и Prunus L. Различия по белковым маркёрам этих и всех остальных таксонов столь велики, что попытки объединить их в единый род Prunus (слива) не являются целесообразными. В роде Prunus в течение третичного периода вымерли в Евразии сначала древнейшие узколистные виды (некоторые из них, мигрировав в Северную Америку через древнюю Североатлантическую сушу, сохранились), затем широколистные виды. К числу древних видов сливы, возникших в тургайских лесах Евразии, относится тёрн (P spinosa L.). Попытки считать его очень молодым гибридным видом [2] — ошибочны. Древним потомком Prunus является род Louiseania Carr. Виды сливы из Восточной Азии и многие североамериканские — молодые виды (рис. 1А).
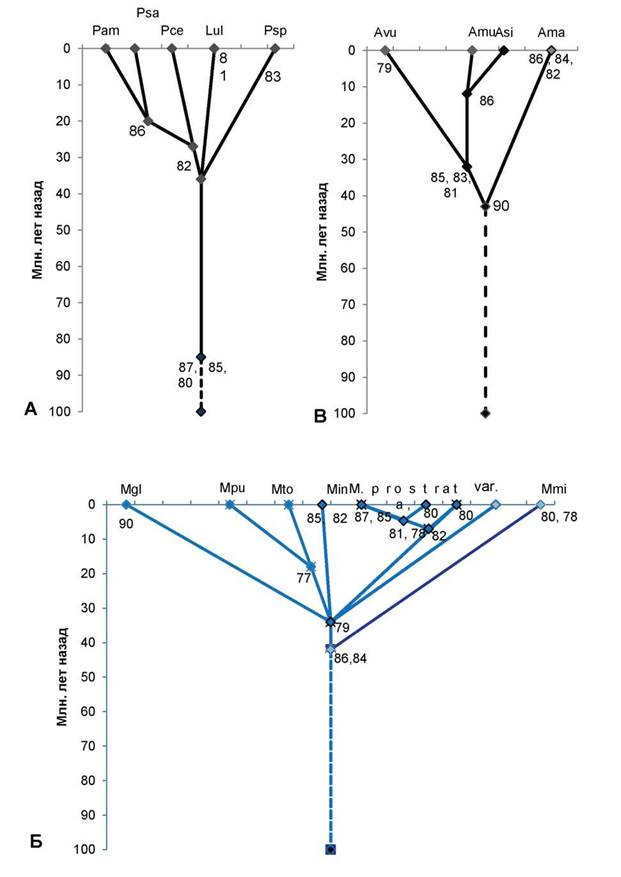
Рис. 1. Схема таксоногенеза внутри родов подсемейства Prunoideae Focke по основным 12S-полипептидам запасных белков-глобулинов семян
Условные обозначения:
А — род слива (Prunus L.): Pam — слива американская (Prunus americana Marsh.); Psa — слива ивовидная, или китайская (Prunus salicina Lindl.); Pce — слива вишневидная, или алыча (Prunus cerasifera Ehrh.); Psp — слива колючая, или тёрн (Prunus spinosa L.); Lul — луизеания (афлатуния) вязолистная [Lou- iseania ulmifolia (Franch.) Pachom.], или Aflatunia ulmifolia (Franch.) Vass. Оба рода выделяются общими древними полипептидными компонентами 87, 85, 80, алыча, сливы ивовидная и американская — молодым
компонентом 82, последние 2 вида сливы — также компонентом 86. Древний тёрн выделяется компонентом 83, а древняя луизеания вязолистная выделяется компонентом 81.
Б — род микровишня (Microcerasus Webb): Mgl — микровишня железистая [Microcerasus glandu- losa (Thunb.) M. Roem.]; Mpu — микровишня низкая [Microcerasus pumila (L.) Erem. et Yushev]; Mto — микровишня войлочная [Microcerasus tomentosa (Thunb.) Erem. et Yushev]; микровишня седая разновидность Блиновского [Microcerasus incana var. blinovskii (Totsch.) Erem. et Yushev]; далее слева направо: микровишня простёртая разновидность двусторонняя [Microcerasus prostrata var. bifrons (Fritsch) Erem. et Yushev], микровишня простёртая разновидность тяньшанская [Microcerasus prostrata var. tianschanica (Po- jark.) Erem. et Yushev], микровишня простёртая разновидность бородавчатая [Microcerasus prostrata var. verrucosa (Franch.) Erem. et Yushev], представленная двумя популяциями — слева типа микровишни красноплодной, справа — типичная микровишня бородавчатая; микровишня мелкоплодная [Microcerasus microcarpa (C. A. Mey.) Erem. et Yushev]. Древние общие компоненты 86, 84, моложе — общий компонент 79. Микровишня железистая выделяется древним компонентом 90, микровишни низкая и войлочная — более молодым компонентом 77, микровишня седая — компонентами 85, 82. Микровишня двусторонняя имеет компоненты 87, 85, тяньшанская — компонент 80 (обе они объединяются компонентами 81, 78), микровишня бородавчатая (типа красноплодной) имеет компонент 80 (объединяясь с предыдущими двумя разновидностями компонентом 82), микровишня бородавчатая (типичная) — самая древняя разновидность, микровишня мелкоплодная — древняя, имеет компоненты 80, 78.
В — род абрикос (Armeniaca Scop.): Avu — абрикос обыкновенный (Armeniaca vulgaris Lam.); Amu — абрикос муме (Armeniaca mume Sieb.); Asi — абрикос сибирский [Armeniaca sibirica (L.) Lam.]; Ama — абрикос маньчжурский [Armeniaca mandshurica (Maxim.) Skvortz.]. Общий древний компонент — 90, абрикосы обыкновенный, муме, сибирский выделяются компонентами 85, 83, 81, из них последние 2 вида — компонентом 86, древний абрикос маньчжурский имеет компоненты 86, 84 и 82.
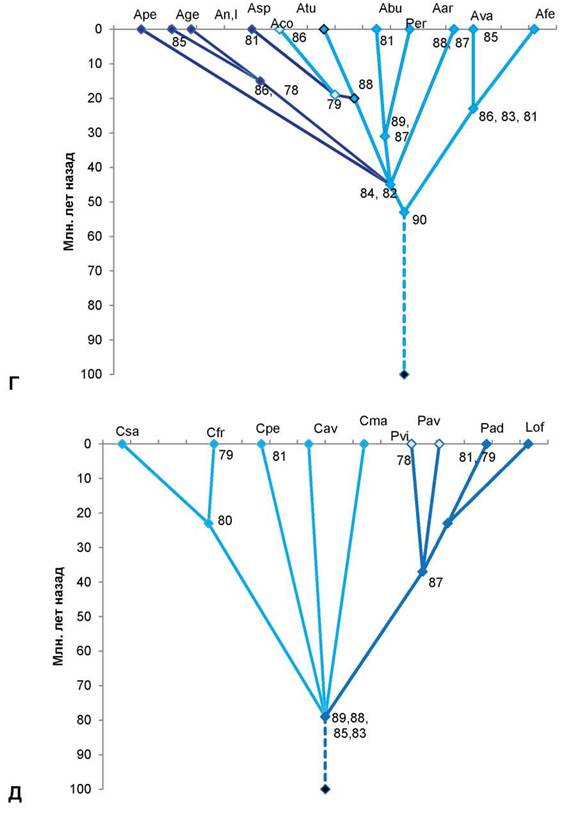
Г — род миндаль (Amygdalus L.): древний компонент 90 дал начало двум ветвям рода — малая ветвь (справа) и большая ветвь (слева). В правой ветви — 2 вида с общими древними компонентами 86, 83, 81, один из нихAfe — миндаль Фенцля [Amygdalusfenzliana (Fritsh) Lipsky]; другой Ava — миндаль Вавилова (Amygdalus vavilovii M. Pop) с компонентом 85. В левой ветви общими древними компонентами являются компоненты 84, 82. Они дали начало Ape — миндалю Петунникова (Amygdalus petunnicovii Litv.); Age — миндалю грузинскому (Amygdalus georgica Desf); An — миндалю низкому (Amygdalus nana L.) и очень близкому к нему Al — миндалю Ледебура (Amygdalus ledebouriana Schlecht.). Последние 3 вида миндаля имеют общий компонент 86, а миндаль грузинский выделяется компонентом 85. Asp—миндаль колючейший (Amygdalus spinosisssima Bunge) с компонентом 81; Atu — миндаль туркменский (Amygdalus turcomanica Lincz.) с компонентом 86, оба миндаля имеют общий компонент 79; Aco — миндаль обыкновенный (Amygdalus communis L.) имеет с миндалями туркменским и колючейшим общий древний компонент 88. Древние компоненты 89, 87 дали начало Abu — миндалю бухарскому (Amygdalus bucharica Korsh.) с компонентом 81 и Aar — миндалю арабскому (Amygdalus arabica Olivier); компонентами 88, 87 выделяется древний род Per — персик (Persica Mill.).
Д — роды вишня (Cerasus Mill.), черёмуха (Padus Mill.), паделлус (Padellus Vass.), лавровишня (Lau- rocerasus Duham.) с общими древними компонентами 89, 88, 85, 83. Csa — вишня сахалинская [Cerasus sachalinensis (Fr. Schmidt) Kom.]; компонентом 79 выделяется Cfr — вишня кустарниковая, или степная (Cerasus fruticosa Pall.), оба этих вида объединяет компонент 80; Cpe — вишня пенсильванская [Cerasus pennsylvanica (L.f.) Loisel.] с компонентом 81; Cav — вишня птичья, или черешня [Cerasus avium (L.) Mo- ench]; Cma — вишня Максимовича [Cerasus maximowiczii (Rupr.) Kom.]; роды с общим компонентом 87: Pav — черёмуха птичья (Padus avium Mill.) с компонентом 78; Pvi — черёмуха виргинская [Padus virginiana (L.) M. Roem.]; Pad — паделлус, магалебка, или антипка [Padellus mahaleb (L.) Vass.] с компонентами 81, 79; Lof— лавровишня лекарственная (Laurocerasus officinalisM. Roem.). Номера в узлах схемы — основные полипептидные электрофоретические компоненты 12S-глобулинов. По вертикальной оси нанесена (в миллионах лет назад) общепринятая в геологии магнитохронологическая шкала [8], где отрезок времени до 65 млн. лет назад — конец мелового периода; 65—54 млн. лет назад — эпоха палеоцена, 54—38 млн. лет назад — эпоха эоцена, 38—26 млн. лет назад — эпоха олигоцена, 26—8 млн. лет назад — эпоха миоцена (третичный период), 8—2 млн. лет назад — эпоха плиоцена; 1,5—2 млн. лет назад и до наших дней — эпоха плейстоцена (четвертичный период). Сплошные линии на схеме — установленные эволюционные связи, прерывистые линии — связи с неизвестными предками.
В роде Cerasus на территории Евразии, Северной Америки растут древнейшие виды (типа черешни, вишен Максимовича, пенсильванской). Но вишня сахалинская, как и степная, лесостепная вишня кустарниковая, являются молодыми видами. Род Padus возник от какого-то вымершего рода вишни во вторичном (урало-западносибирском)
центре происхождения вишен, а роды Padellus (магалебка) и особенно род Lauroceresus, продолжающие линию древних вишен, — наиболее молодые роды (рис. 1Д).
Остальные роды сливовых более молодые, возникли в эпохи палеогена. В роде Amyg- dalus (миндаль) издревле развивались две линии, из которых древнейшим был предок миндалей с бороздчатым эндокарпием (типа миндаля Вавилова, Фенцля и др.). Мнение о культигенном и гибридном происхождении миндаля Вавилова [2] — также ошибочное [3]. К древним миндалям второй линии относятся миндали, растущие в степной зоне (в том числе A. nana L.), их предок возник одновременно с родом Persica (персик). Персик расселялся в лесах Евразии, вымер, сохранившись в основном на востоке Азии. Из миндалей этой второй линии очень древним является миндаль Петунникова, который сохранился в горах на севере Средней Азии. Виды миндаля из районов Передней Азии, юга Средней Азии — сравнительно молодые, но старше северных степных миндалей (рис. 1Г).
Из рода микровишня (ошибочно относимого к вишне) древнейшими являются виды из западной части Евразии, в особенности микровишня мелкоплодная, имеющая резко реликтовый ареал в Передней Азии. Виды востока Евразии, Северной Америки — более молодые, но сохранили в себе признаки древних лесных видов Евразии (рис. 1Б). В Средней, особенно в Передней Азии между видами микровишни идут интенсивные процессы гибридизации, приводящие к взаимной интрогрессии части их геномов, но на автополиплоидной основе [3].
Много спорного было в возрасте и истории рода Armeniaca (абрикос). Древнейшим видом является абрикос маньчжурский с реликтовым типом ареала. Более молодой вид — абрикос обыкновенный, самые молодые — абрикос сибирский (с крупнейшим ареалом) и абрикос муме (или китайский) с ареалом на юге Восточной Азии (рис. 1А).
Общего предка подсемейства Prunoideae установить пока сложно. Не изучены по белковым маркёрам род Emplectocladus из Северной Америки, ряд родов, близких к вишне, черёмухе и растущих в Восточной Азии. Однако метод белковых маркёров показывает, что центром происхождения родов сливовых являются древние Тургайская и Древнесредиземноморская области Евразии в пределах современной западной (а не восточной) части Евразии [3, 4]. Учитывая, что древнейшими таксонообразующими белковыми маркёрами у двудольных растений являются низкомолекулярные полипептиды основных 128-глобулинов [5, 6], можно полагать, что предок был ближе к роду Cerasus. При формировании подсемейства сливовых самостоятельной была линия, давшая начало родам слива (с луизеанией), вишня (с черёмухой, паделлусом, лавровишней). В другой линии сливовых выделились миндали древнейшего происхождения (типа миндаля Вавилова), затем только предковая линия дивергировала на «абрикосовую» и предка родов микровишня, персик и остальных видов миндаля (рис. 2).
Таким образом, род миндаль имеет сборное происхождение, и это мнение часто высказывалось в литературе. Интересно, что данные по белковой эволюции подтверждают данные по анатомии листа сливовых [9] в части того, что род абрикос, хотя и внешне близок к сливе (и с нею частично гибридизирует), но родственен древней «миндальной» ветви.
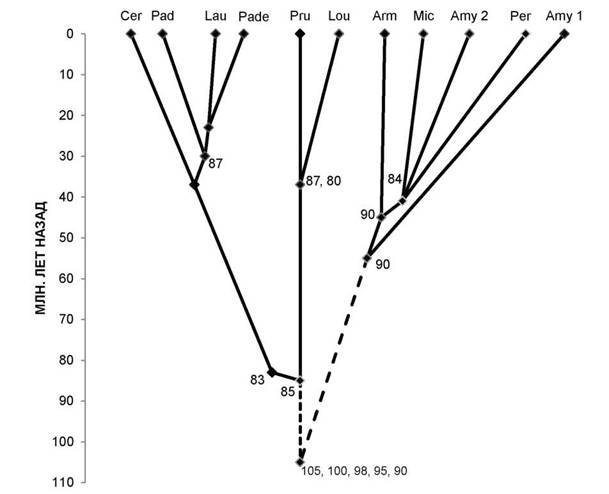
Рис. 2. Схема таксоногенеза родов подсемейства Prunoideae Focke по 12S-полипептидам. Условные обозначения: Cer — род Cerasus Mill. (вишня); Pad — PadusMill. (черёмуха); Lau — Laurocerasus Duham. (лавровишня); Pade — Padellus (L.) M. Roem. (паделлус, магалебка); Pru — Prunus L. (слива); Lou — Lou- iseania Carr, (луизеания), или Aflatunia Vass. (афлатуния); Arm — Armeniaca Scop. (абрикос); Mic — Micro- cerasus Webb (микровишня); Amy 2 — виды Amygdalus L. (миндаль, большая ветвь, см. рис. 1); Amy 1 — то же, но малая ветвь; Per — Persica Mill. (персик). Клада «вишнеслив» маркируется древним компонентом 85, при этом «вишнёвые» роды — компонентом 83, а роды слива и луизеания — молодыми компонентами 87 и 80 (компонент 87 маркирует также паделлус, черёмуху, лавровишню). Клада «абрикос, микровишня, персик, миндаль» возникла много позже на основе древнего компонента 90, а позднее — более молодого компонента 84.
На рисунках 1 и 2 существенно уточнён возраст ряда таксонов, известный в литературе (см. [7, с. 92]). Цифрами указаны полипептидные компоненты разной степени древности; сплошные линии — установленные эволюционные связи, прерывистые — связи с неизвестными предками. Остальные обозначения аналогичны рисунку 1.
Список литературы
Кимура М. Молекулярная эволюция: теория нейтральности. М. : Мир, 1985. 400 с.
Ерёмин Г. В. Отдалённая гибридизация косточковых плодовых растений. М. : Колос, 1985. 280 с.
Авдеев В. И. Плодовые растения Средней Азии, их происхождение, классификация, исходный материал для селекции : дис. ... д-ра с.-х. наук. СПб. : ВНИИР им. Н. И. Вавилова, 1997. 328 с.
Авдеев В. И. К истории происхождения видов подсемейства сливовых (Rosaceae) в степной зоне // Тр. Ин-та биоресурсов и прикладной экологии. Оренбург : ОГПУ, 2000. Вып. 1. С. 45—58.
Авдеев В. И. Этапы формирования степных ландшафтов в Евразии. Аспекты эволюции видов Poa- ceae // Известия ОГАУ 2009. № 2. С. 59—65.
Авдеев В. И., Саудабаева А. Ж. Сравнительный анализ адаптаций по полипептидным маркёрам у абрикосов Оренбуржья // Приёмы повышения адаптивности косточковых культур, вопросы осеверения и расширения границ садоводства : сб. материалов междунар. симпозиума. Челябинск : НПО «Сад и огород», 2011. С. 45—51.
Благовещенский А. В., Александрова Е. Г. Биохимические основы филогении высших растений. М. : Наука, 1974. 104 с.
Сузюмов А. Е. Земной магнетизм и тектоника дна океанов // Новое в жизни, науке, технике. М. : Знание, 1981. № 1. 48 с.
Соколова Е. А. Значение анатомических признаков для систематики представителей подсемейства Prunoideae (Rosaceae) : автореф. дис. ... д-ра биол. наук. СПб. : ВНИИР им. Н. И. Вавилова, 2000. 28 с.
Похожие работы
-
Виды аминокислот
Аминокислоты являются основными структурными единицами молекул белковых веществ. При гидролизе белков различной природы всегда получают смесь 20 аминокислот.
-
Сущность антропосоциогенеза
Фундаментальным отличием стада животных от человеческого общества является наличие в человеческом обществе формируемых сознанием людей социальных законов.
-
Генетическая история человечества
Эволюция и филогенетическое древо. Эволюция и ДНК. Люди генетически близки друг к другу. Предки человека. ДНК и восстановление истории человечества. «Митохондриальная Ева» и «Y-хромосомный Адам». Эволюция популяций и рас.
-
Гены и история
Этнографы и генетики давно ищут способы подобраться к изучению истории происхождения различных этнических групп. Можно только приветствовать работы учёных нового поколения, которые оперируют генетическими маркёрами различных этнических групп.
-
Фауна и экология листоедов подсемейства galeruciane (coleoptera: chrysomelidae] Белгородской области
На территории Белгородской области выявлено 17 видов листоедов галеруцин (Coleoptera: Chrysomelidae: Galerucinae). Среди них пищевые предпочтения не выяснены для одного вида.
-
Современные очаги происхождения северного абрикоса в Евразии
В северной зоне садоводства на территории Евразии в настоящее время существуют 7 различных очагов происхождения местного сортимента абрикоса. По каждому из очагов кратко приведён их селекционный потенциал.
-
Белковые маркёры у ряда культивируемых злаков в Оренбуржье
Методом электрофореза запасных белков семян изучены 6 сортов ячменя, 7 сортов пшеницы и 2 ранее не исследованных образца сорного и культивируемого проса. Все сорта пшеницы, ячменя хорошо идентифицируются с помощью белковых маркёров. Образцы проса сорного и культивируемого по маркёрам наиболее близки к пшенице.
-
Биохронологические методы
Определение возраста по годичным кольцам, или дендрологический метод, дает абсолютные данные за последние 3000 лет истории Земли. Они основываются на том, что деревья ежегодно откладывают кольца роста, благодаря которым утолщаются стволы.
-
Эволюционная классификация
Подобно эмпирическим схемам классификации, эволюционная классификация основана на простом факте, что в природе встречаются ясно отличающиеся друг от друга группы, такие, как птицы, пингвины, летучие мыши, жуки и т. п.
-
Белки семян как маркеры в решении проблем генетических ресурсов растений, селекции и семеноводства
Обсуждаются основные направления и перспективы использования белковых и ДНК-маркеров в решении прикладных и теоретических проблем генетических ресурсов растений, селекции, сортоиспытания, семеноводства и семенного контроля.