Название: Физиологические эффекты гормонов, плохо проникающих в клетку
Вид работы: учебное пособие
Рубрика: Биология
Размер файла: 148.61 Kb
Скачать файл: referat.me-19133.docx
Краткое описание работы: Действие гормонов на клеточном уровне. Плохо проникающие в клетку гормоны (белково-пептидные, катехоламины), их действие через рецепторы на клеточной мембране. Использование аденилатциклазной, фосфоинозитидной, гуанилатциклазной и тирозинкиназной систем.
Физиологические эффекты гормонов, плохо проникающих в клетку
Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), соединяются с рецепторами на клеточной мембране и вызывают внутриклеточные эффекты через вторые посредники.
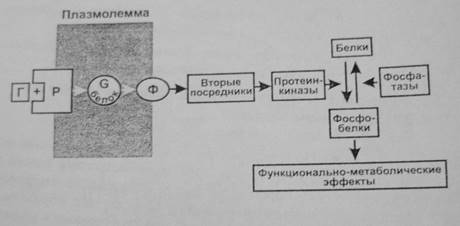
Большинство гормонов после присоединения к рецепторам активируют регулируемый гуанилнуклеотидами G-белок мембраны. Активация состоит в обмене ГДФ на ГТФ в α-субъединице G-белка. При этом происходит его распад на α-субъединицу и βγ-димер, имеющие самостоятельные регуляторные эффекты. Более изучено действие α-субъединицы, являющейся ферментом ГТФазой. Гидролиз ГТФ до ГДФ освобождает энергию, используемую для передачи регуляторного сигнала. ГДФ-α-субъединица соединяется с βγ-димером, вновь образуя ГДФ—G-белок. Имеется несколько видов G-белков (GS , Gi ,Gq ,GO ), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (GS ), так и ингибировать (Gi ) образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.
В действии гормонов, плохо проникающих в клетки, можно выделить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатциклазную и тирозинкиназную.
Аденилатциклазная система. Эту систему используют гормоны, осуществляющие свой эффект через аденилатциклазу и синтез второго посредника—циклического АМФ (3,5-цАМФ). Действие гормона может, как активировать, так и тормозить его образование.
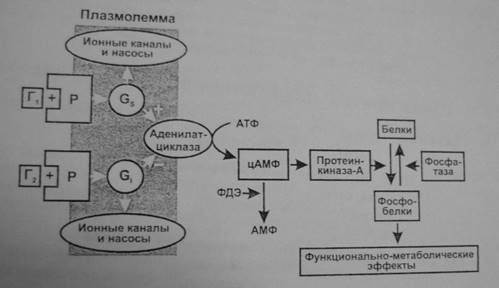
- Комплекс «гормон + рецептор» активирует GS –белок, который активирует аденилатциклазу и образование цАМФ (например, так действуют адреналин через β1 /β2 -рецепторы, АДГ через V2 -рецепторы, глюкагон, АКТГ, ТТГ, ФСГ, ЛГ, МСГ, кортиколиберин, соматолиберин, кальцитонин, хориогонадотропин человека, паратгормон).
- Комплекс «гормон + рецептор» активирует Gi -белок, который тормозит аденилатциклазу и образование цАМФ (например, так действуют адреналин через α2 -рецепторы, ангиотензин II через АТ-рецепторы, аденозин через А1 -рецепторы, соматостатин, опиоиды через μ-, δ- и κ-рецепторы, ацетилхолин через М2 -холинорецепторы гладкомышечных клеток).
- цАМФ влияет на функции клетки через изменение активности протеинфосфокиназы-А, фосфорилирующей различные клеточные белки: мембранные (изменяется ее проницаемость), ферменты (изменяется метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение трансляции), микротрубочек (двигательные эффекты, изменение формы клетки) и др.
Фосфоинозитидная система. Эту систему используют гормоны, которые, действуя через Gq -белок, активируют в мембране фосфолипазу С.
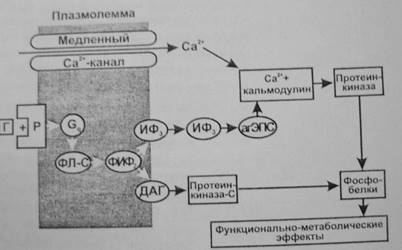
Она образует из фосфолипида мембраны (фосфатидилинозитолдифосфата) два посредника—инозитол—1,4,5—трифосфат (ИФ3 ) и 1,2—диацилглицерол (ДАГ).
- ИФ3 , переходя в цитоплазму, действует на рецепторы гладкой ЭПС, что приводит к открыванию Са2+ -каналов и выходу Са2+ в цитозоль. Са2+ при этом выступает в качестве третьего посредника, который связывается с белком кальмодулином и активирует Са2+ -кальмодулинзависимую протеинкиназу, фосфорилирующую эффекторные белки, влияющие на функции клеток, их пролиферацию и дифференцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфолипазы—А2 и синтез простагландинов, тромбоксана, простациклина, гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной железы, гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов). (Роль Са2+ как посредника действия гормонов обусловлена его сильным влиянием при низких концентрациях 10-8 —10-6 М на метаболизм и функции клетки. Гормоны могут увеличивать концентрацию Са2+ в цитозоле как в результате открывания Са2+ -каналов клеточной мембраны, так выхода Са2+ из гладкой ЭПС. В клетке есть специальные белки, связывающие Са2+ при низкой концентрации—кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2+ к этим белкам изменяет их конфигурацию и свойства. Это приводит к изменению многочисленных функций клеток—метаболической, секреторной, сократительной, биоэлектрической и др.)
- ДАГ, оставаясь на внутренней поверхности мембраны, активирует протеинкиназу С, фосфорилирующую эффекторные белки (например, активирует киназу легкой цепи миозина), а также синтез простагландинов из арахидоновой кислоты, что изменяет функции клетки.
- Через ИФ3 и ДАГ действуют адреналин (α-рецепторы), вазопрессин (V1 ), ацетилхолин (М1 ), ангиотензин II (AT1 ), гистамин (Н1 ), серотонин (5-НТ2 ), холецистокинин, окситоцин, ВИП и др.
Гуанилатциклазная система. Некоторые гормоны (например, ацетилхолин, цитокины) действуют через гуанилатциклазу, образующую второй посредник—циклический ГМФ (цГМФ). Гуанилатциклаза существует в двух формах—мембраносвязанной и растворимой в цитозоле.
- Мембраносвязанная гуанилатциклаза является частью рецептора плазмолеммы, ее активация гормоном (например, Na+ -уретическим) увеличивает образования цГМФ, который стимулирует протеинкиназу G, фосфорилирующую белки.
- Растворимая в цитозоле гуанилатциклаза содержит в своем составе гемм, с которым связывается сигнальная молекула – NO (оксид азота образуется из аргининав ряде клеток—эндотелии, нейронах и др. – под действием ацетилхолина, механических раздражителей и др.). Активированная оксидом азота гуанилатциклаза через протеинкиназу G фосфорилирует белки (например, так действует ацетилхолин, вызывая расширение сосудов).
Тирозинкиназная система характеризуется особыми рецепторами, которые путем фосфорилирования и дефосфорилирования сигнальных белков по тирозиновым остаткам, регулируют процессы роста, дифференцировки, выживания, метаболизма и смерти клеток. Эту систему используют некоторые гормоны—инсулин, инсулиноподобные факторы роста (или соматомедины), факторы роста эпидермиса, тромбоцитов и др. Многие рецепторы к таким гормонам содержат в своём составе тирозинкиназу, которая при действии гормона аутофосфорилирует внутреннюю часть рецептора по остаткам тирозина. Активированный таким образом рецептор передает через сложный и недостаточно изученный комплекс посредников фосфорилирующее и дефосфорилирующее влияние по остаткам тирозина на многие сигнальные молекулы цитоплазмы и ядра. Фосфорилирование и дефосфорилирование белков может приводить как к их активации, так и инактивации.
Похожие работы
-
Растительная клетка, ее строение
Растительная клетка, ее строение Типичная растительная клетка содержит хлоропласты и вакуоли и окружена целлюлозной клеточной стенкой. Плазматическая мембрана (плазмалемма), окружающая растительную клетку, состоит из двух слоев липидов и встроенных в них молекул белков. Молекулы липидов имеют полярные гидрофильные «головки» и неполярные гидрофобные «хвосты».
-
Принципы гормональной регуляции
План: Принципы гормональной регуляции (Гормоны, группы гормонов, влияние гормонов) Системные принципы гормональной регуляции физиологических функций
-
Роль гормонов в регуляции кальция и фосфатов парагормон кальцитриол кальцитонин
Московская Государственная академия тонкой химической технологии им. M.В. Ломоносова Реферат по курсу: Регуляция клеточной активности Тема: Роль гормонов в регуляции кальция и фосфатов (парагормон, кальцитриол, кальцитонин)
-
Функциональная система. Гормональная регуляция функций
Теория функциональной системы П. Анохина. Узлы и компоненты функциональной системы. Афферентный и эфферентный сигналы. Гормональная регуляция функций. Гипоталамо-гипофизарная система. Тканевые гормоны. Гормоны вилочковой железы. Энкефалины и эндорфины.
-
Гормоны
Химическая природа и классификация гормонов. Биороль простагландинов и тромбоксанов. Регуляция секреции гормонов. Гормональная регуляция углеводного, липидного, белкового и водно-солевого обмена. Роль циклазной системы в механизме действия гормонов.
-
Эндокринная система животных и человека
Роль эндокринной системы в регуляции основных процессов жизнедеятельности животных и человека. Свойства, классификация, функции, и биологическая роль гормонов эндокринных желез. Анализ проблемы йоддефицитных заболеваний человека и животных в России.
-
Физиология желез внутренней секреции животных
Железы внутренней секреции у животных. Механизм действия гормонов и их свойства. Функции гипоталамуса, гипофиза, эпифиза, зобной и щитовидной железы, надпочечников. Островковый аппарат поджелудочной железы. Яичники, желтое тело, плацента, семенники.
-
Кортикостероиды
Биохимическое исследование кортикостероидов как подкласса стероидных гормонов, производимых исключительной корой надпочечников. Химическое строение и минералокортикоидная активность кортикостероидов. Анализ биологической роли и биосинтез кортикостероидов.
-
Железы внутренней секреции
Виды гипофиза, их структура и функции в организме. Состав тканей и назначение надпочечников. Условия проявления функциональных эффектов адреналина. Деятельность эндокринных тканей в органах, обладающих неэндокринными функциями. Значение плаценты, тимуса.
-
Нейротрансмиттеры и головной мозг
Общая характеристика нейротрансмиттеров. Нейромедиаторы. Нейромодуляторы.