Название: Двойное оплодотворение
Вид работы: реферат
Рубрика: Биология
Размер файла: 39.38 Kb
Скачать файл: referat.me-19507.docx
Краткое описание работы: Двойное оплодотворение - половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Особенности протекания процессов микроспорогенеза и мегаспорогенеза. Понятие и роль опыления.
Двойное оплодотворение
, половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon ) и рябчике (Fritillaria orientalis ). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез , а также опыление .
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно .
Пыльцевое зерно снаружи покрыто двумя оболочками: экзиной и интиной . Экзина – верхняя оболочка более толстая и пропитана спороленнином – жироподобным веществом. Это позволяет пыльце выдерживать существенные температурные и химические воздействия. В экзине находятся проростковые поры, до опыления закрытые «пробочками». Интина содержит целлюлозу и эластична. В пыльцевом зерне имеются две клетки: вегетативная и генеративная .
Мегаспорогенез осуществляется в семязачатке . Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок . Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка – яйцеклетка . На халазальном полюсе располагаются антиподы . Таким образом, зрелый зародышевыый мешок содержит 7 клеток.
Опыление заключается в переносе пыльцы с тычинок на рыльце пестика.
Оплодотворение . Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте – так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия. Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, образуя зиготу, которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Таким образом, эндосперм у покрытосеменных триплоидный и вторичный , т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения . Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
У голосеменных в оплодотворении участвует один спермий и эндосперм возникает до оплодотворения в результате деления мегаспоры, т.е. он гаплоидный и первичный .
Антиподы и синергиды растворяются после оплодотворения, а интегументы преобразуются в семенную кожуру.
Апомиксис – развитие зародыша из неоплодотворенной клетки. Формы апомиксиса зависят от того, из каких частей семязачатка развивается зародыш. При партеногенезе (партенос- девственный) зародыш возникает из неоплодотворенной яйцеклетки. Если зародыш развивается из какой-либо другой клетки гаметофита (антиподы, синергиды), то такой процесс называется апогамия . В случае образования зародыша из клеток нуцеллуса, интегументов, не входящих в гаметофит, говорят об апоспории . Апомиксис распространен среди эволюционно развитых групп растений. За счет апомиксиса растения не зависят от опыляющих агентов
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ
половой процесс у покрытосеменных растений, при к-ром оплодотворяются как яйцеклетка, так и центр. клетка зародышевого мешка.
Д. о. открыл рус. учёный С. Г. Навашин
в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis)
. В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия
сливается с ядром яйцеклетки,
ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш,
из центр. клетки - эндосперм.
В зародышевых мешках с трёхклеточным лицевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид,
которая при этом разрушается (в ней видны остатки ядра синер-гиды и вегетативного ядра пыльцевой трубки)
; вторая синергида впоследствии отмирает. Далее оба спермия вместе с изменённой цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центр. клеткой. Затем спермин разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром,
другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним,а иногда и с обоими полярными ядрами. Спермин теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермин в виде неизменённых клеток наблюдаются и в 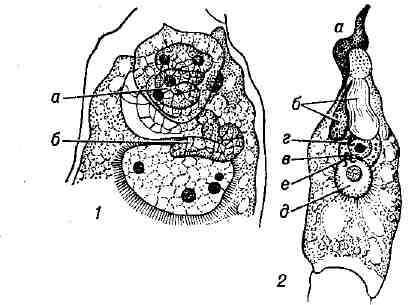
зародышевом мешке.
Двойное оплодотворение; 1 - у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) - с одним из полярных ядер (второе полярное ядро не изображено); 2 - у подсолнечника; а - пыльцевая трубка; 6 - синергиды (одна из них повреждена пыльцевой трубкой); в - яйцеклетка; г - спермий в контакте с ядром яйцеклетки; д - центральная клетка; е - второй спермий в контакте со вторичным ядром зародышевого мешка.
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние к-рых могут вырьировать. У скерды и нек-рых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у мн. растений они удлинённые, иногда извитые, б. или м. хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Ге-расимова-Навашина) различать два типа Д. о.: премитотическое -ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе) ; постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце к-рой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоид-но. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центр. клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплои-ден. Следствие Д. о. - Ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает неск. пыльцевых трубок, спермий первой из них участвуют в Д. о., спермии остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в "Известиях Петербургской Академии Наук".
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных.
Двойное оплодотворение - отличительный признак, отделяющий покрытосеменных от голосеменных.
Открытие С. Г. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке.
Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
Похожие работы
-
Бесполое размножение организмов
• При бесполом размножении потомки развиваются из одной материнской клетки или группы клеток (части материнского организма). • По наследуемым свойствам образующиеся потомки идентичны материнскому организму и называются клонами.
-
Партеногенез
(Parthenogenesis - от греч. parthenos - девушка, девственница + genesis-зарождение) - форма полового размножения, при котором развитие организма происходит из женской половой клетки (яйцеклетки) без оплодотворения ее мужской (сперматозоид).
-
Таблица по биологии амебы и прочие твари
Тип Простейшие. Класс Саркодовые.Амеба. Бактерия Вакуоль пищеварительная вакуоль [KL1] сократительная вакуоль Выделять- что? куда? Выделить-с помощью чего?
-
Морфология и анатомия цветка
Цветок как специальный орган размножения, характерный для цветковых растений. Строение цветка, его осевые части и боковые придатки. Сосудистый скелет цветка. Причины упрощения в строении пучков. Функции и строение околоцветника, тычинок и пестика.
-
Семенные растения и их общая характеристика
Особенности моноспорического типа образования женского гаметофита цветковых растений. Нуцеллус как развитая паренхиматическая ткань, которая занимает центральную часть семяпочки. Эволюционное развитие семенных растений. Мужской гаметофит у голосеменных.
-
Характеристика голосеменных растений
История появления голосеменных растений и их современные представители. Особенности цикла развития и состав водопроводящей системы; принципы оплодотворения. Характеристика класса хвойных, их значение в биосфере и в хозяйственной деятельности человека.
-
Строение цветкового растения: корень, цветок и плод
Изучение вегетативных органов растений. Их видоизменения (колючка, усик, клубни, луковицы), функции и строение. Цветки и соцветия - генеративные органы растения. Описание процесса опыления и оплодотворения растений. Распространение плодов и семян.
-
Развитие зародыша человека
Министерство образования Российской Федерации Управление образования администрации города Чебоксары МОУ «Кадетская Школа» Реферат на тему: Развитие зародыша человека
-
Ароморфозы растений и животных (WinWord 98)
#ТЕМА: Ароморфозы растений и животных #РАЗДЕЛ: Биология #НАЗНАЧЕНИЕ: таблица #ФОРМАТ:WinWord 98 #АВТОР: Александра М. #СДАВАЛСЯ: школа № 606 г. Санкт-Петербург. учитель Корнилова О.В
-
Гинецей растений
Характеристика, значение и строение гинецея. Простой и сложный гинецей. Положение завязи в цветке. Структура семязачатка до оплодотворения. Происхождение и эволюция гинецея. Плацентация и её типы, расположение плацент. Завязь, строение и типы семязачатка.